

Неврохимична основа на шизофренията
- Допаминовата хипотеза продължава да бъде валидна невробиологична теория за шизофренията1,2.
- Допаминовите рецептори имат различно разпределение и афинитет.
- Преполага се, че позитивните симптоми при шизофрения са свързани с хиперактивност на допаминовите рецептори в мезолимбичната система, а хипоактивността в мезокортикалната система е свързана с проявата на негативни и когнитивни симптоми.
В този раздел:
Допаминова хипотеза
Допаминовата хипотеза за шизофренията се е превърнала в теория въз основа на косвени доказателства, от проведени клинични наблюдения след антипсихотично лечение. Образните изследвания позволяват по-добро разбиране на подлежащите патофизиологични механизми3.
Допаминът и мозъкът
Допаминът има специфично действие върху различните подтипове рецептори, които са отговорни за различни функции (Префронтална област на мозъчната кора: Когнитивна функция5, Мотивация/възнаграждение6–10, Негативни симптоми при шизофрения11, Дорзален стриатум: Двигателен контрол12, болест на Паркинсон1, Дистония1, Вентрален стриатум: Мотивация/възнаграждение6–10, Позитивни симптоми при шизофрения11,13, Биполярно разстройство14, Депресия8, Зависимости15, ХАДВ16), нарушението на тези процеси може да доведе до множество симптоми, свързани с психични заболявания. Психотропните лекарства, които влияят върху допаминовите рецептори, могат да подобрят много от тези симптоми, така че е необходимо по-добро разбиране на допаминовата функция в различните области на мозъка и механизмите, които са в основата на шизофренията4.
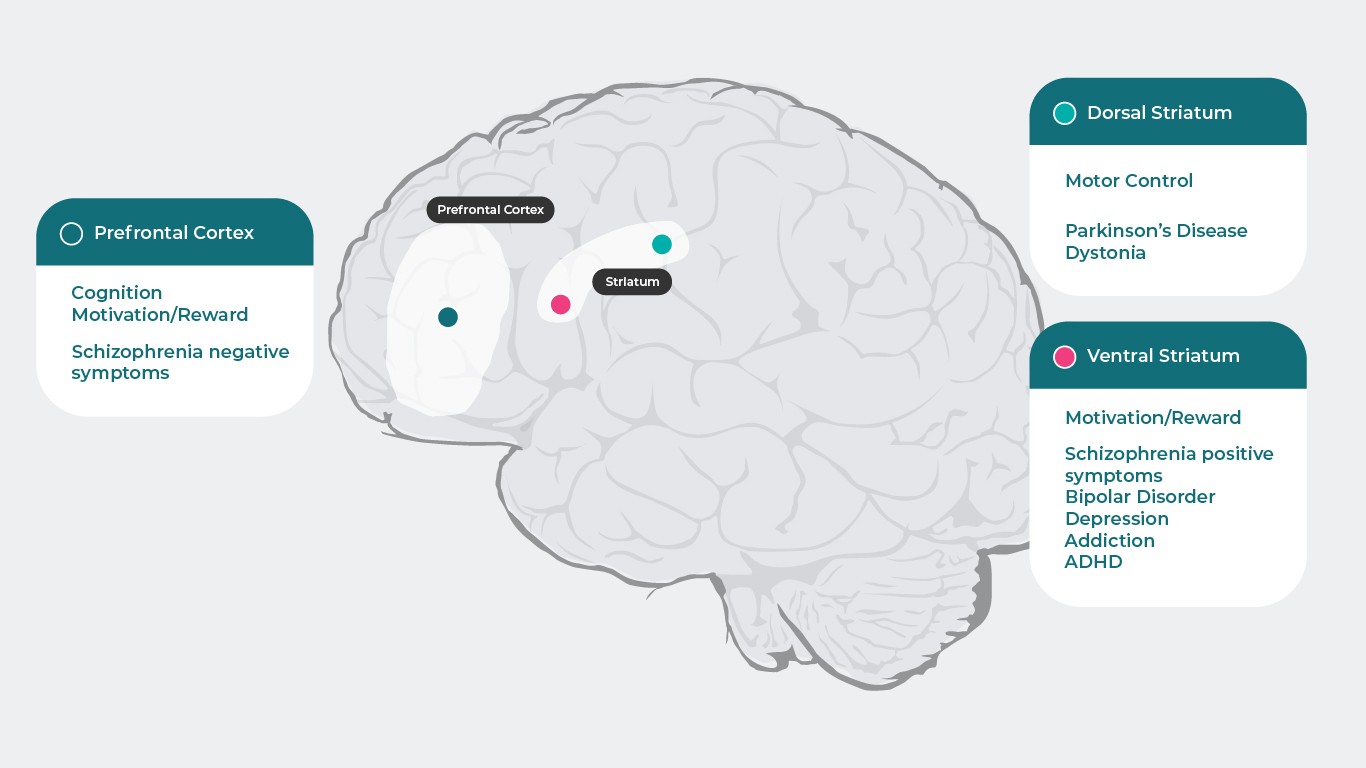
Източници: Адаптирано по Stahl SM. Essential Pharmacology 4th edition 20131 ; Cools R, D’Esposito M. Biol Psychiatry. 2011;69(12):e113-125.5; Hamid AA, et al. Nat Neurosci. 2016;19(1):117-126.6; Koob GF, et al. NIDA Res Monogr. 1994;145:1–187; Biesdorf C, et al. Neurobiol Learn Mem. 2015;123:125-1398; Der-Avakian A, et al. Trends Neurosci. 2012 Jan;35(1):68-77.9; Leggio GM, et al. Eur Neuropsychopharmacol. 2008 Apr;18(4):271-7.10; Stahl SM. Prim Care Companion J Clin Psychiatry.2003;5(3)9-1311; Plowman EK, et al. J Parkinsons Dis. 2011;1(1):93-10012; Dichter GS, et al. J Neurodev Disord. 2012;4:19513; Ashok AH, et al. Mol Psychiatry.2017 May;22(5):666–67914; Orio L, et al. Addict Biol. 2010;15(3):312-32315; Volkow ND, et al. J Neurosci. 2012;32(3):841-84916.
Допаминови рецептори: подтипове
Има 5 типа допаминови рецептори, които могат да бъдат разделени на 2 основни класа. D1-подобните рецептори включва допаминовите D1 и D5 рецептори, D2-подобните рецептори включват D2, D3, и D4 рецептори. От клинична гледна точка се приема, че допаминовите D2, D3 и D1 рецептори като цяло имат най-голямо отношение към патофизиологията на шизофренията. В зависимост от рецепторния подтип допаминовите рецептори може да проявяват различни ефекти в една и съща сигнална система – да инхибират или да стимулират4. Например D1 рецепторите могат да стимулират cAMP, а D2 и D3 рецепторите могат да инхибират тази вторична сигнална молекула.
По този начин допаминовите D1, D2 и D3 рецептори могат да изпълняват много специфични, но различни невробиологични функции при наличието на допамин, което позволява един невротрансмитер да контролира различни физиологични, поведенчески и патологични състояния4,17.
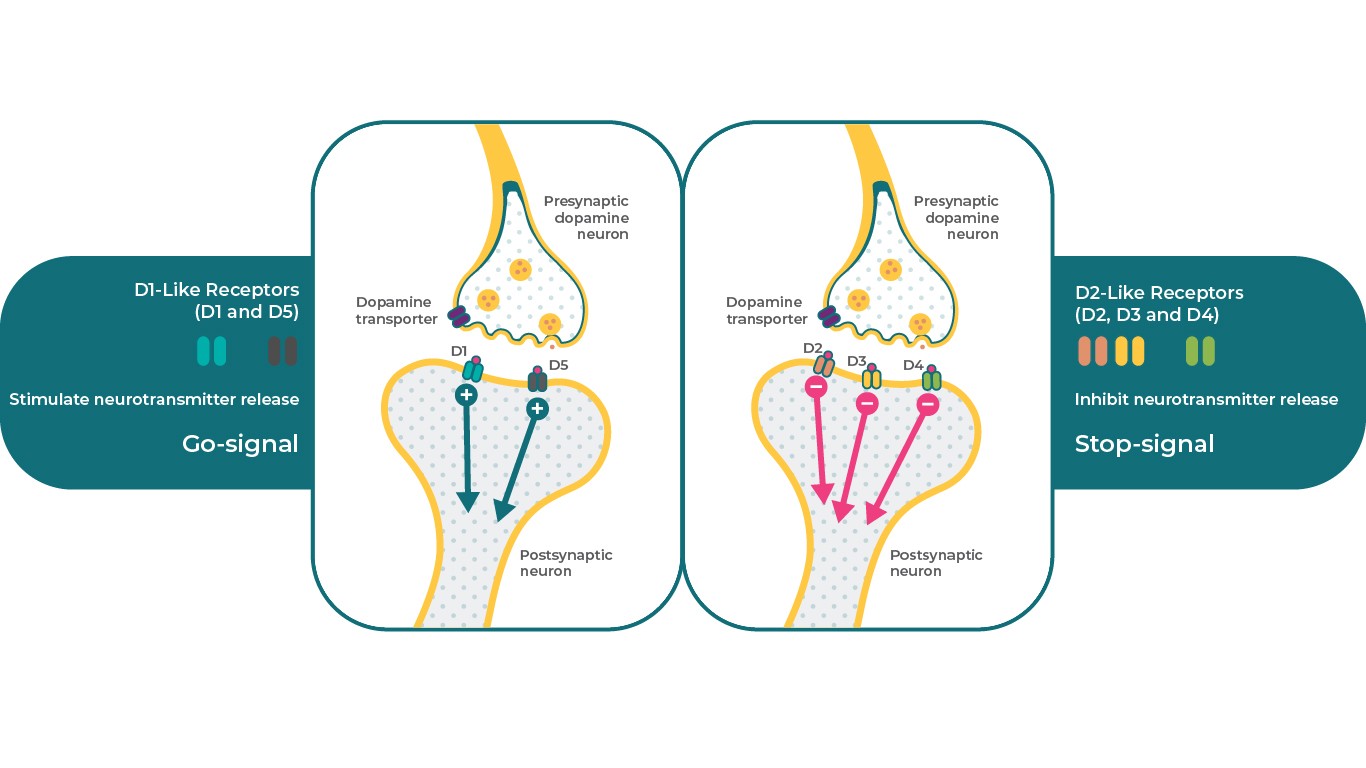
Характеристики на допаминовите D1-подобни и D2-подобни рецептори в мозъка.
Източници: Адаптирано по Stahl, S. M. Dazzled by the dominions of dopamine: Clinical roles of D3, D2, and D1 receptors. CNS Spectr. 22, 305–311 (2017)4 Източници: Адаптирано по References: Adapted from Stahl, S. M. Dazzled by the dominions of dopamine: Clinical roles of D3, D2, and D1 receptors. CNS Spectr. 22, 305–311 (2017)4
Допаминови рецептори: афинитет
Всеки подтип допаминови рецептори има различен афинитет към допамин. Въз основа на клинични данни PDSP Ki, D3 рецепторите19 имат най-голям афинитет към допамин, следвани от D219,20, D421 и D518 рецепторите със среден афинитет и D118,19 рецепторите са с най-малък афинитет към допамин4.
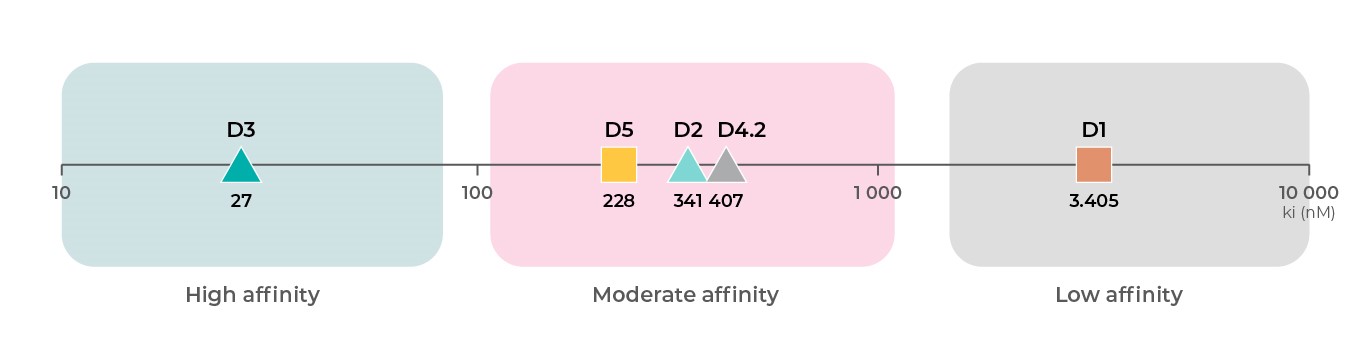
Източници: Всички данни за свързването са от базата данни за Ki на PDSP.
Affinity to D1 subtype: Toll L, NIDA Res Monogr 1998, 178:44046619 ; Sunahara RK, Nature 1991, 350:614-61918
Affinity to D2 subtype: Michaelides MR 1995, J Med Chem 38:3445-344720; Toll L, NIDA Res Monogr 1998, 178:44046619
Affinity to D3 subtype: Toll L, NIDA Res Monogr 1998, 178:44046619; Cussac D, Naunyn Schmied Arch Pharmacol 2000, 361:569-572;
Affinity to D4 subtype: Tallman JF, J Pharm Exp Ther 1997, 282:1011-101921;
Affinity to D5 subtype: Sunahara RK, Nature 1991, 350:614-61918; Stahl SM, CNS Spect 2016, 21:123-12722.
Допаминови рецептори: разпределение4,22-27
Разпределението на допаминовите рецептори в мозъка също е различно при отделните подтипове. Например се предполага, че свръхактивното освобождаване на допамин към D2 постсинаптичните рецептори във вентралния стриатум причинява позитивни симптоми на психоза. Поради това допаминовите D2 рецептори са главна мишена за повечето антипсихотици, използвани за лечение на шизофрения. D3 рецепторите са силно експресирани в лимбичната област, хипоталамуса и вентралната тегментална област/субстанция нигра, области, за които се смята, че участват в регулацията на когнитивната функция, настроението и мотивацията. В префронталния кортекс, където почти няма експресия на допаминови D2 и D3 рецептори, допаминовата невротрансмисия се контролира в по-голяма степен от постсинаптичните D1 рецептори.
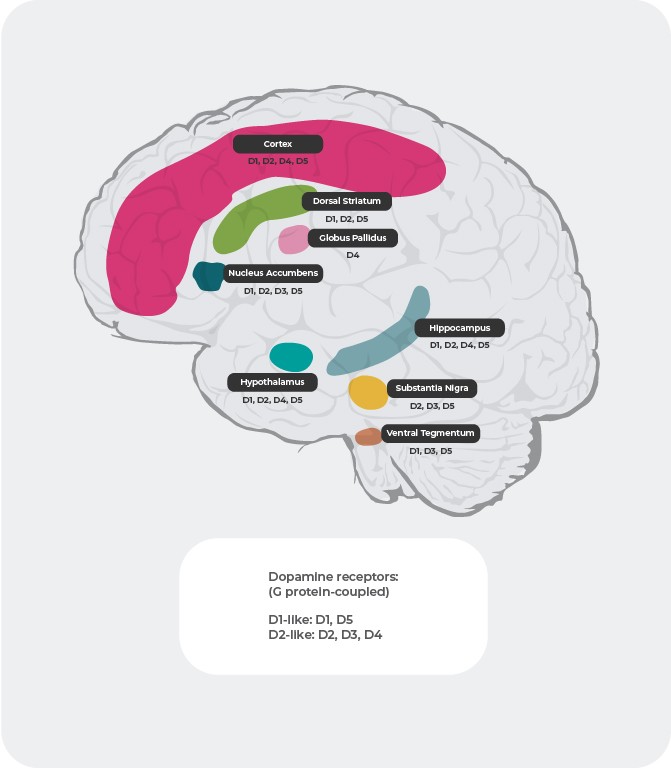
Източници: Адаптирано по Stahl, CNS spec. 2017;22:305-311 18; Fatemi SH. The medical basis of psychiatry. 4th edition. Sringer 2016;p93-9427
Допаминергични пътища
В човешкия мозък има 4 главни допаминергични пътища: мезолимбичен, мезокортикален, нигростриатален и тубероинфундибуларен път27:
Мезолимбичен път: мезолимбичният път, наричан също път на възнаграждението, предава допамин от вентралната тегментална област (ВТО) към вентралния стриатум. Вентралната тегментална област е разположена в средния мозък, а вентралния стриатум – в предния мозък, като включва нуклеус акумбенс и обонятелния туберкул.
Мезокортикален път: Мезокортикалният път, за който се смята, че участва в когнитивната функция и емоциите, предава допамин от вентралната тегментална област към префронталната област на мозъчната кора.
Нигростриатален път: Нигростриаталният път, който участва в регулацията на движенията, предава допамин от компактната част на субстанция нигра (SNc) в средния мозък към нуклеус каудатус и путамен в дорзалния стриатум.
Тубероинфундибуларен път: тубероинфундибуларният път предава допамин от инфундибуларното ядро на хипоталамуса към хипофизата. Този път регулира секрецията на хормони от хипофизата, включително пролактин.
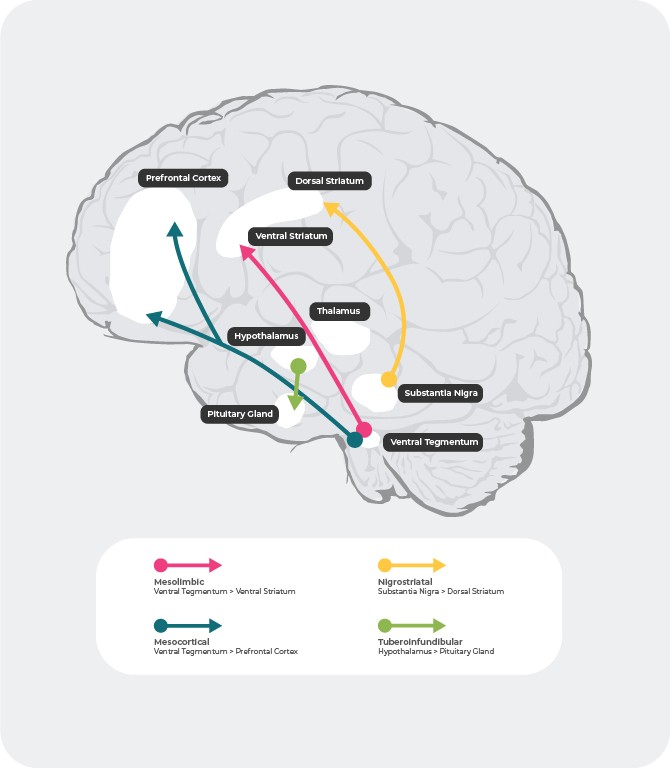
Източници: Адаптирано по Fatemi SH. The medical basis of psychiatry. 4th edition. Sringer 2016;p93-9427
Допамин и шизофрения28,29
Допаминовата хипотеза за шизофрения предполага, че хиперактивната допаминова невротрансмисия в субкортикалната и лимбичната зона на мозъка е свързана с позитивните симптоми на шизофрения30, а хипофункцията на мезокортикалните допаминови проекции към префронталната област на мозъчната кора може да отговаря за негативните и когнитивните симптоми31,32.
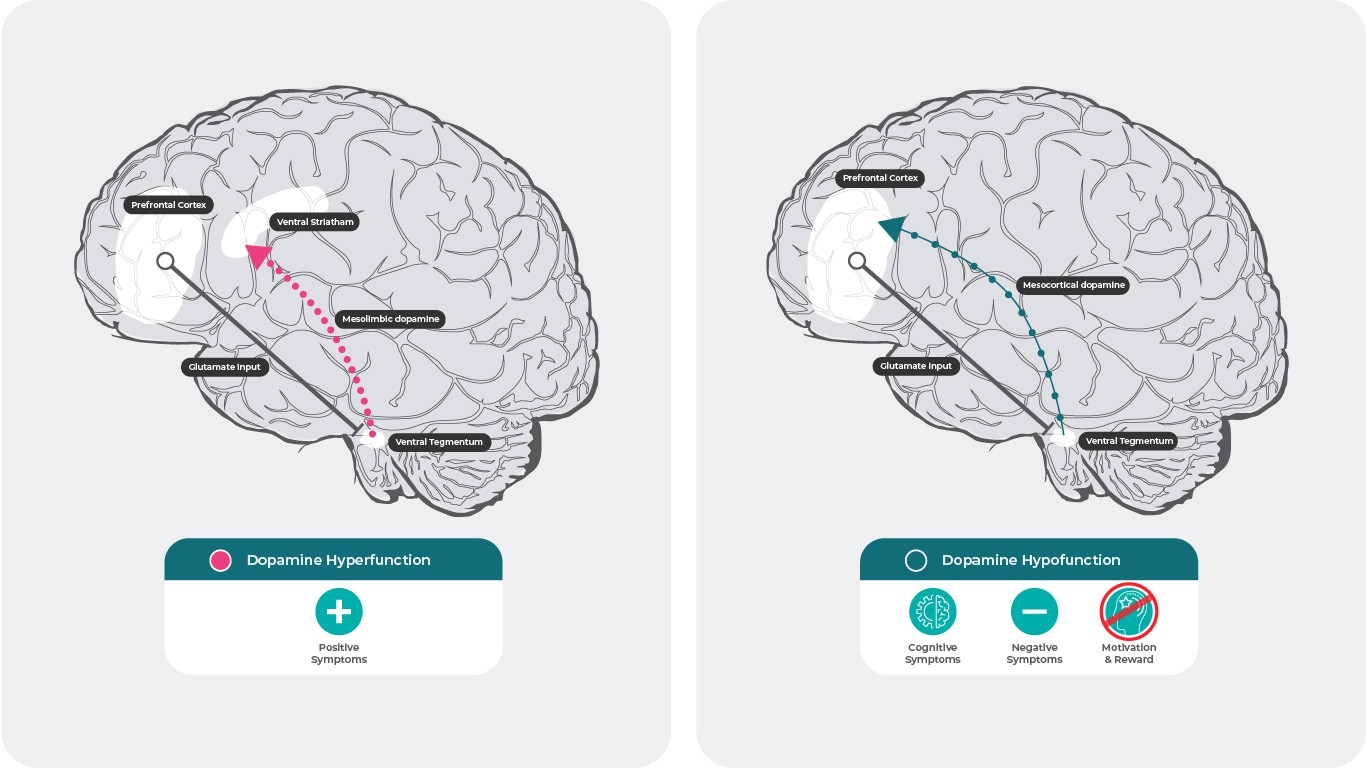
Ефект на допаминовата хиперфункция и хипофункция в мозъка.
Източници: Адаптирано по Stahl SM. 4th ed. New York, NY: Cambridge University Press; 20131; Howes J Psychopharmacol. 2015 February; 29(2): 97–115.; 35. Elert E, Nature 2014. 508. s2-s329
Предложени са няколко теории за допаминовата дисрегулация, които да обяснят патофизиологията на шизофренията33,34. Например в проучване се предполага, че патофизиологията при шизофрения може да бъде причинена главно от дисрегулация на глутамата, водеща до допаминергичен дисбаланс35.
Каскадата започва с възбуда на глутаматните неврони, които изпращат електрически импулс (1), водещ до освобождаване на глутамат от синапса (2).
При нормални условия след това глутаматът се свързва с инхибиторните GABA неврони и възбудните глутаматни неврони. При пациентите с шизофрения обаче има нарушение на свързването на глутамат с GABA невроните (3), което води до неосвобождаване на инхибиторни невротрансмитери (4). Глутаматът обаче се свързва с възбудните глутаматни неврони и при липса на инхибиторни GABA неврони (5) се изпраща усилен сигнал към мозъчния ствол (6).
Като последица се освобождава твърде много глутамат от синапса, който свръхстимулира допаминовите неврони и други инхибиторни GABA неврони (7).
Свръхстимулираните GABA неврони инхибират прекомерно различен допаминов неврон, като по този начин потискат активността му (8). Този допаминов неврон с потисната активност предава интермитентни сигнали от вентралната тегментална област към префронталната област на мозъчната кора, където се освобождава твърде малко допамин (9), което води до хиподопаминергично състояние в мезокортикалната система, предизвикващо проявата на негативни симптоми1.
Впоследствие тези свръхстимулирани допаминови неврони предават усилени сигнали от вентралната тегментална област към вентралния стриатум, което е причина за прекомерно освобождаване на допамин (10), като резултатът е хипердопаминергия в мезолимбичната система и позитивни симптоми.
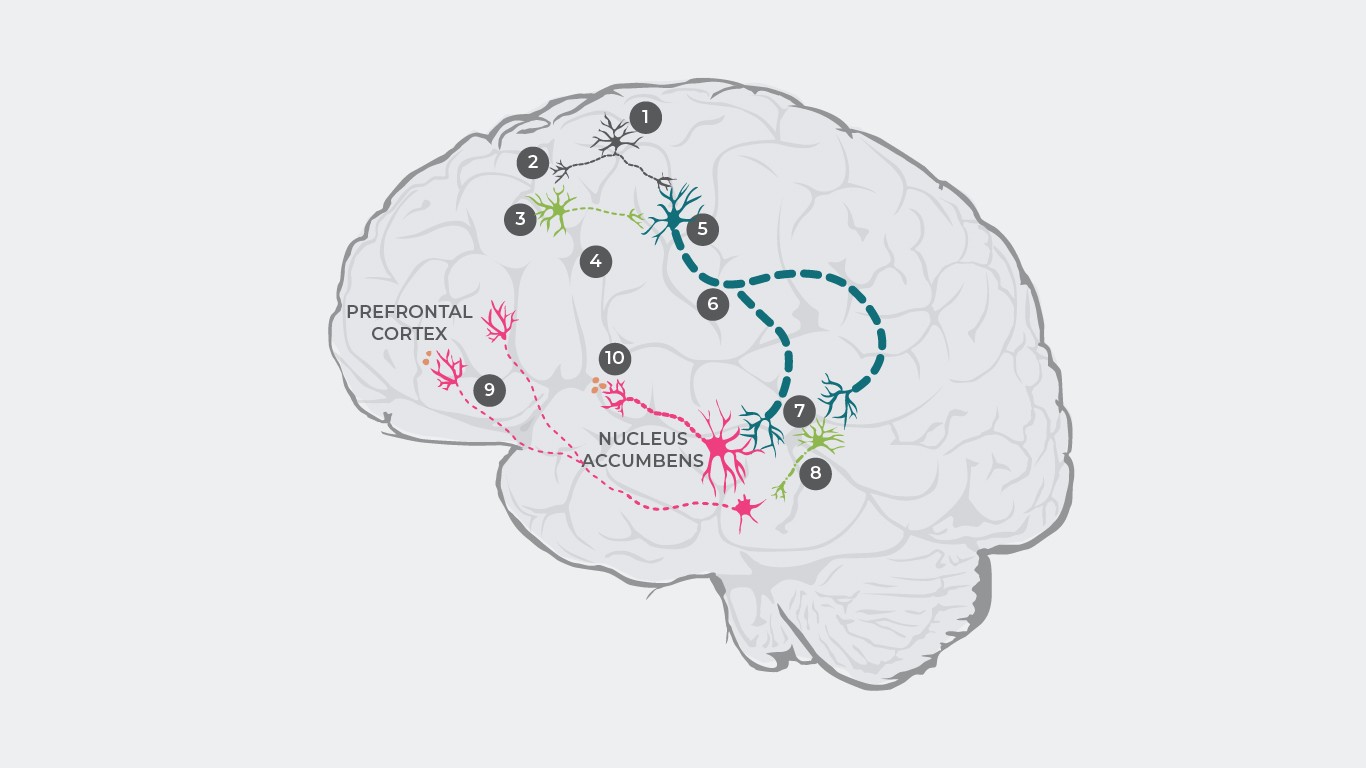
Източници: Адаптирано по Stahl SM. 4th ed. New York, NY: Cambridge University Press; 20131; Howes J Psychopharmacol. 2015 February; 29(2): 97–11529; Elert Nature volume 508, pages S2–S3 (03 April 2014)35;
Проучването подкрепя тезата, че повишената невротрансмисия на допаминовите D2 рецептори в субкортикалната и лимбичната зона в мозъка допринася за позитивните симптоми31,32. Не са ясни обаче механизмите, които причиняват негативни и когнитивни симптоми.
Понастоящем се предполага, че пониженото освобождаване на допамин в префронталната област на мозъчната кора се причинява от активиране на пресинаптични допаминови D3 авторецептори във вентралната тегментална област, които достигат до префронталната област на мозъчната кора. От подтиповете допаминови рецептори D3 рецепторите са с най-висок афинитет към допамина. За разлика от D1 или D2 рецепторите, D3 рецепторите могат да бъдат стимулирани в условия на ниски нива на освобождаване на допамин, което може да повлияе на флуктуиращите ефекти от взривообразното фазово освобождаване на допамин. Блокирането на това понижение спира инхибирането на освобождаването на допамин и засилва невротрансмисията. Това повишение на нивото на допамина може да неутрализира хиподопаминергичното състояние и да доведе до подобрение на негативните симптоми и когнитивната функция посредством активиране на D1 рецепторите в префронталната област на мозъчната кора, които не са активни при ниски нива на допамин32.
При животински модели, включително проучвания при примати, се постигат най-благоприятни когнитивни ефекти при оптимизиране на активността на D1 рецепторите в префронталната област на мозъчната кора. Твърде силната и твърде слабата допаминова активност на кортикалните D1 рецептори е свързана с когнитивна дисфункция. Хипотетично лекарства, които блокират или свръхстимулират D1 рецепторите, могат да нарушат регулацията на допаминовата стимулация и да окажат влияние върху когнитивните и негативните симптоми и симптомите, свързани с настроението, като нарушат хармонията на невротрансмисията в кортикалните синапси. Също така е възможно антагонистите на D3 рецепторите да спрат инхибирането на освобождаването на допамина и да хармонизират кортикалникте кръгове, което от своя страна може да доведе до подобрена регулация на когнитивната функция, настроението и негативните симптоми4.
Източници
- Stahl, S. M. Stahl’s Essential Psychopharmacology: Neuroscientific Basis and Practical Applications. (Cambridge University Press, 2013).
- Seeman, P. Schizophrenia and dopamine receptors. Eur. Neuropsychopharmacol. 23, 999–1009 (2013).
- Kaar, S. J., Natesan, S., McCutcheon, R. & Howes, O. D. Antipsychotics: Mechanisms underlying clinical response and side-effects and novel treatment approaches based on pathophysiology. Neuropharmacology (2019). doi:10.1016/j.neuropharm.2019.107704
- Stahl, S. M. Dazzled by the dominions of dopamine: Clinical roles of D3, D2, and D1 receptors. CNS Spectr. 22, 305–311 (2017).
- Cools, R. & D’Esposito, M. Inverted-U-shaped dopamine actions on human working memory and cognitive control. Biol. Psychiatry 69, e113-125 (2011).
- Hamid, A. A. et al. Mesolimbic dopamine signals the value of work. Nat. Neurosci. 19, 117-126. (2016).
- Koob, G. F., Caine, B., Markou, A., Pulvirenti, L. & Weiss, F. Role for the mesocortical dopamine system in the motivating effects of cocaine. NIDA Res. Monogr. Ser. 145, 1-18. (1994).
- Biesdorf, C. et al. Dopamine in the nucleus accumbens core, but not shell, increases during signaled food reward and decreases during delayed extinction. Neurobiol. Learn. Mem. 123, 125–139 (2015).
- Der-Avakian, A. & Markou, A. The neurobiology of anhedonia and other reward-related deficits. Trends Neurosci. 35, 68–77 (2012).
- Leggio, G. M., Micale, V. & Drago, F. Increased sensitivity to antidepressants of D3 dopamine receptor-deficient mice in the forced swim test (FST). Eur. Neuropsychopharmacol. 18, 271–277 (2008).
- Stahl, S. M. Describing an atypical antipsychotic: receptor binding and its role in pathophysiology. Prim. Care Companion J. Clin. Psychiatry 5, 9–13 (2003).
- Plowman, E. K., Thomas, N. J. & Kleim, J. A. Striatal dopamine depletion induces forelimb motor impairments and disrupts forelimb movement representations within the motor cortex. J. Parkinsons. Dis. 1, 93–100 (2011).
- Dichter, G. S., Damiano, C. A. & Allen, J. A. Reward circuitry dysfunction in psychiatric and neurodevelopmental disorders and genetic syndromes: Animal models and clinical findings. J. Neurodev. Disord. 4, 19 (2012).
- Ashok, A. H. et al. The dopamine hypothesis of bipolar affective disorder: The state of the art and implications for treatment. Mol. Psychiatry 22, 666–679 (2017).
- Orio, L., Wee, S., Newman, A. H., Pulvirenti, L. & Koob, G. F. The dopamine D3 receptor partial agonist CJB090 and antagonist PG01037 decrease progressive ratio responding for methamphetamine in rats with extended-access. Addict. Biol. 15, 312–323 (2010).
- Volkow, N. D. et al. Methylphenidate-elicited dopamine increases in ventral striatum are associated with long-term symptom improvement in adults with attention deficit hyperactivity disorder. J. Neurosci. 32, 841–849 (2012).
- Beaulieu, J. M., Espinoza, S. & Gainetdinov, R. R. Dopamine receptors – IUPHAR review 13. Br. J. Pharmacol. 172, 1–23 (2015).
- Sunahara, R. K. et al. Cloning of the gene for a human dopamine D5 receptor with higher affinity for dopamine than D1. Nature 350, 614–619 (1991).
- Toll, L. et al. Standard binding and functional assays related to medications development division testing for potential cocaine and opiate narcotic treatment medications. NIDA Res. Monogr 178, 440–466 (1998).
- Michaelides, M. R. et al. (5aR, 11bS)-4,5,5a,6,7, 11b-hexahydro-2-propyl-3-thia-5-azacyclopent-1-ena[c]-phenanthrene-9,10-diol (A-86929): A potent and selective dopamine D1 agonist that maintains behavioral efficacy following repeated administration and characterization of its dia. J. Med. Chem. 38, 3445–3447 (1995).
- Tallman, J. F. et al. Ngd 94-1 – identification of a noval, high-affinity antagonist at the human dopamine D-4 receptor 1. J. Pharmacol Exp Ther 282, 1011–1019 (1997).
- Stahl, S. M. Mechanism of action of cariprazine. CNS Spectr. 21, 123–127 (2016).
- Gurevich, E. V. & Joyce, J. N. Distribution of dopamine D3 receptor expressing neurons in the human forebrain comparison with D2 receptor expressing neurons. Neuropsychopharmacology 20, 60–80 (1999).
- Mengod, G. et al. Visualization of dopamine D1, D2 and D3 receptor mRNA’s in human and rat brain. Neurochem. Int. 20, Suppl:33S-43S (1992).
- Meador-Woodruff, J. H. et al. Dopamine receptor mRNA expression in human striatum and neocortex. Neuropsychopharmacology 15, 17–29 (1996).
- Fuxe, K. et al. Dopaminergic Systems in the Brain and Pituitary. in Basic and Clinical Aspects of Neuroscience (eds. Flückinger, E., Müller, E. E. & Thorner, M. O.) (Springer, 1985). doi:10.1007/978-3-642-69948-1_2
- Fatemi, S. H. & Clayton, P. J. The medical basis of psychiatry: Fourth edition. The Medical Basis of Psychiatry: Fourth Edition (2016). doi:10.1007/978-1-4939-2528-5
- Howes, O. D. & Kapur, S. The dopamine hypothesis of schizophrenia: Version III – The final common pathway. Schizophr. Bull. 35, 549–562 (2009).
- Howes, O., McCutcheon, R. & Stone, J. Glutamate and dopamine in schizophrenia: An update for the 21st century. J. Psychopharmacol. 29, 97–115 (2015).
- Desbonnet, L. Modeling the Psychopathological Dimensions of Schizophrenia. in Handbook of Behavioural Neuroscience 267–284 (2016).
- Toda, M. & Abi-Dargham, A. Dopamine hypothesis of schizophrenia: Making sense of it all. Curr. Psychiatry Rep. 9, 329–336 (2007).
- Stahl, S. M. Drugs for psychosis and mood: Unique actions at D3, D2, and D1 dopamine receptor subtypes. CNS Spectr. 22, 375–384 (2017).
- Grace, A. A. & Gomes, F. V. The circuitry of dopamine system regulation and its disruption in schizophrenia: Insights into treatment and prevention. Schizophr. Bull. 45, 148–157 (2019).
- Grace, A. A. Dysregulation of the dopamine system in the pathophysiology of schizophrenia and depression. Nat. Rev. Neurosci. 17, 524–532 (2016).
- Elert, E. Aetiology: Searching for schizophrenia’s roots. Nature 508, S2-3 (2014).
CNS Spectrums
Dazzled by the dominions of dopamine: clinical roles of D3, D2, and D1 receptors.
антипсихотици, РАЗЛИЧНИ МЕХАНИЗМИ... антипсихотици, РАЗЛИЧНИ МЕХАНИЗМИ...
(COD: 300020/R05. Представено в AIFA 16/04/2020) Изглежда, че клиничният профил на безопасност на ефикасност на типичните антипсихотици зависи от високия Изглежда, че клиничният профил на безопасност на ефикасност на типичните антипсихотици зависи от високия афинитет към и пълната антагонистична активност спрямо
повече…КАК ДЕЙСТВА REAGILA? КАК ДЕЙСТВА НАШИЯТ ПРОДУКТ?
(COD: 300020/R06. Представено в AIFA 16/04/2020) Cariprazine има висок афинитет към допаминовите D3 и D2 рецептори, както и към серотониновите 5 HT2B и 5 HT1A, Научете повече за механизма на действие
повече…
